
Immunotherapy has entered a new era, and B-cells, long considered secondary to T cells, are now recognised as central to both disease and therapy. Beyond the well-established antibody production function, B-cells have direct effector function, are potent antigen-presenting cells, shape T cell responses, and exert profound regulatory functions. The broader role of B-cells in cancer, infectious disease, and autoimmunity is becoming well understood, and the potential to harness B-cell multi-functional therapeutic benefits is rapidly coming into focus; the future path for translational development is paramount and unlocks an opportunity for a new arm of therapy.
The challenge lies in the transition from blunt interventions, such as broad B-cell depletion, to precision approaches that leverage specific B-cell subsets. Both adoptive transfer of engineered B-cells and in vivo manipulation are advancing, with antigen-specific strategies emerging as the most promising way to balance efficacy with safety (Figure 1).
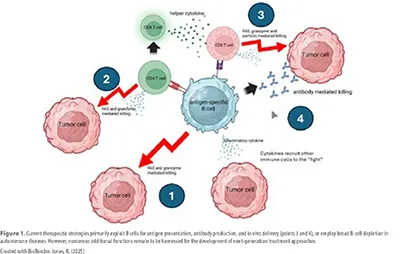
A Historical Timeline: The evolution of B-cell targeting
1990s–2000s: Anti-CD20 antibodies (rituximab, ocrelizumab) revolutionised the treatment of B-cell lymphomas and multiple B-cell-related autoimmune diseases. These early successes validated B-cell surface proteins as effective therapeutic targets to deplete all B-cell modalities, revealing unintended consequences of indiscriminate B-cell depletion, including hypogammaglobulinemia, recurrent infections, and loss of protective immunity. For example, repeated rituximab courses required for maintenance of response, combined with higher glucocorticoid doses in ANCA-associated vasculitis, increased the risk of hypogammaglobulinemia (very low IgG) at 6 months in ~43% of treated patients; lower IgG levels were associated with serious infections.
In children with autoimmune diseases, hypogammaglobulinemia after rituximab was frequent, particularly in CNS disease or vasculitis, and correlated with serious infections.
2010s: B-cells gained recognition in cancer biology, with tumor-infiltrating B-cells (TIL-Bs) and tertiary lymphoid structures (TLS) linked to improved survival in melanoma, lung cancer, and breast cancer. In melanoma, baseline high T cell/low B-cell gene signatures were associated with better overall survival in a trial of dabrafenib + trametinib. Meanwhile, recognition of memory B-cells has refined our understanding of durable vaccine responses, which may impact new vaccine development.
2020s: Emerging preclinical studies of engineered B-cells demonstrate the feasibility of adoptive transfer for antibody delivery and antigen presentation. For example, in mice, engineered B-cells expressing broadly neutralizing antibodies (bNAbs) against HIV showed durable secretion, homing to germinal centers, and neutralization activity. Simultaneously, experimental models of autoimmunity suggest that regulatory B-cells (Bregs) could be expanded or transferred to promote tolerance. For instance, ex vivo expanded B10 cells (an IL-10-producing Breg subset) markedly inhibited disease symptoms in experimental autoimmune encephalomyelitis (EAE).
In cancer, Tertiary lymphoid structures (TLS) correlate with response to checkpoint inhibitors, suggesting that reprogramming intratumoral B-cells could enhance current therapies.
2030 and beyond: There is a lot of potential for the development of antigen-specific B-cell therapies in clinical trials spanning oncology, chronic viral infection, and autoimmunity.
Cancer: Toward Precision Activation of Specific B-cells
In cancer, the most important accomplishment has been the recognition that B-cells within tumors are not passive bystanders but active participants in anti-tumor immunity. Their presence often predicts better outcomes, and adoptive transfer of engineered B-cells is showing early therapeutic promise and supports the move from tumor tolerance to immune coordination.
Current accomplishments:
Engineered B-cells in mice secreting tumor-specific antibodies have shown sustained antibody production, coupled with antigen presentation that bolsters T cell responses. For instance, a recent preclinical study engineered B-cells to express an anti-HPV antibody targeting an intracellular tumor-antigen; those B-cells not only secreted antibody but also induced CD4+ and CD8+ T cell activation.
Adoptive B-cell transfer using antigenic engineering has been shown to reduce tumor growth in several mouse models. For example, in the review “Exploiting B-cell Transfer for Cancer Therapy,” engineered B-cells were modified ex vivo (e.g., forced expression of costimulatory ligands, antigen loading) and transferred, resulting in improved immune responses and tumor control.
Current accomplishments: Pre-clinical evidence of antigen-specific B cell potential.
Tumor-associated regulatory B-cells (Bregs) remain a barrier. These subsets secrete immunosuppressive cytokines such as IL-10 and TGF-β, dampening anti-tumor immunity.
Systemic depletion strategies risk eliminating both beneficial and detrimental subsets, including those that contribute to tumor surveillance or protective immunologic memory.
Tumor antigen heterogeneity and immune evasion must be addressed: precise targeting and modulation of antigen-specific B-cells and targeted delivery are key for tipping the balance toward immunity.
This strengthens the rationale for antigen-specific manipulation: programming or expansion of B-cells to respond to tumor antigens, sparing protective compartments such as memory B-cells and non-pathogenic B-cell subsets.
Infectious Disease: Beyond Broad Vaccination
B-cells are the backbone of vaccines based on immunologic antibody responses. Annual vaccination cycles are a symptom of B-cell dysfunction combined with inefficient target decisions, yielding transient responses. The next wave of B-cell development is to direct their therapeutic use. Adoptive transfer of engineered B-cells could provide long-term, renewable antibody production against conserved epitopes in mutable viruses like HIV, influenza, and other viruses—avoiding the need for repeated monoclonal antibody infusions and reducing or eliminating annual development of new or annually updated vaccines.
Current accomplishments:
In vivo engineering of B-cells via adeno-associated virus or “AAV” plus CRISPR/Cas9 has allowed for endogenous B-cell integration of broadly neutralising antibody (bNAb) genes. Engineered B-cells homed to germinal centers and bone marrow, secreted multiple isotypes, and neutralised heterologous viruses.
Transfer of engineered “emAb” B-cells against RSV in mice demonstrated durable protection with phenotypes consistent with both long-lived plasma cells in the bone marrow and switched memory in the spleen.
Precise targeting and modulation of antigen-specific B-cells and targeted delivery are key for tipping the balance toward cancer immunity and must overcome:
Tumor-associated regulatory B-cells (Bregs) in the immunosuppressive tumor microenvironment (TME). These subsets secrete cytokines such as IL-10 and TGF-β, which dampen anti-tumor immunity and T cell function.
Tumor-antigen heterogeneity and immune evasion (precise targeting and modulation of antigen-specific B-cells and targeted delivery are key for tipping the balance toward immunity; moved)
Risk of non-disease-specific B-cell depletion that eliminates both helpful and harmful subsets, including those contributing to tumor surveillance or protective immunologic memory.
Current issues:
Conventional vaccination strategies can fail in the elderly or immunocompromised populations as well as in chronic infection settings, where B-cell exhaustion or antigenic variation reduces antibody quality.
Polyclonal activation of non-disease-specific B-cells can generate off-target or autoreactive responses.
Ensuring the safety of gene editing (off-target effects, vector immunogenicity) and long-term persistence is non-trivial.
These limitations support the shift toward antigen-specific B-cell manipulation, whether through the adoptive transfer of engineered cells or in vivo expansion of targeted clones.
Autoimmunity: From Depletion to Regulation
The autoimmune field has been transformed first by anti-CD20 therapy and more recently with CD19 CAR T cells mediated B cell depletion that also eliminates plasma cells, which continues to validate B-cells as drivers of disease in multiple sclerosis, lupus, and rheumatoid arthritis. The current accomplishment is undeniable: B-cell depletion has provided durable remission for many patients in a select set of disease areas.
However, these therapies are not without cost: Non-disease-specific B cell depletion results in systemic immunosuppression.Long-term depletion can cause serious infections, impair vaccine responses, and eliminate protective or regulatory subsets.
In pediatric autoimmune conditions, rituximab-associated hypogammaglobulinemia is frequent and often persistent, especially in patients with central nervous system disease, and correlates with infections that are more serious.
Secondary hypogammaglobulinemia has also been documented in nephrotic syndrome patients treated with rituximab; persistent hypogammaglobulinemia in ~22% of patients.
These clinical realities highlight the need for the development of antigen-specific approaches. Expanding or engineering regulatory B-cells offers the first truly antigen-specific tolerance strategy.
Adoptive transfer of regulatory B-cells (Bregs) engineered or expanded ex vivo to suppress autoimmune responses while preserving protective compartments. Ex vivo expanded B10 cells have been shown to suppress EAE in mice
In vivo manipulation of Bregs, using cytokines, small molecules, or engineered ligands to selectively expand regulatory subsets at sites of inflammation. Studies have identified IL-21 plus CD40 stimulation as a potent driver of B10 cells in mice, yielding 10^6 fold expansion and disease suppression.
Expanding or engineering regulatory B-cells offers the first truly antigen-specific tolerance strategy—potentially the immune equivalent of gene therapy.
Mixed antigen-peptide chimeric B-cells: In the non-obese diabetic (NOD) mouse model, B lymphocytes treated ex vivo with LPS, electroporated with MHC-peptide constructs (class I or II) linked to autoantigenic peptides, could specifically suppress autoreactive CD8+ or CD4+ T cells and prevent autoimmune diabetes.
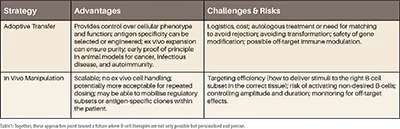
Adoptive Transfer vs. In Vivo Manipulation: Converging Paths
The two dominant strategies in B-cell immunotherapy are adoptive transfer and in vivo manipulation, and are likely to progress in parallel, as each approach has its own advantages and drawbacks.
Outlook
Lessons learned from the T cell field are clear: blunt strategies yield early success, but antigen-specific precision is needed for durable impact. For B-cells, the time has come to move beyond depletion and toward targeted, antigen-specific immunotherapy. Together with the accomplishments of the past decades—anti-CD20 depletion in cancer and autoimmunity, recognition of the role of B-cells in cancer, and proof-of-concept engineering in infectious disease—have laid the groundwork. The side effects of non-specific therapies now highlight the urgent need for antigen-specific B-cell approaches that can preserve protective functions while targeting pathogenic ones.
Looking forward, it is reasonable to expect over the next decade:
First-in-human studies using engineered B-cells in adoptive transfer approaches for infectious disease and cancer.
Initial clinical trials of in vivo manipulation to target and expand or suppress specific B-cell subsets in autoimmunity.
Thus, the next decade may not just broaden the scope of immunotherapy—it may redefine it, centered on what could become the ‘B-cell Decade.
Thus, the next decade may not just broaden the scope of immunotherapy—it may redefine it, with B-cells at the center as the “B-cell Decade”.
References
1. Khojah, A.M., Miller, M.L., Klein-Gitelman, M.S., Curran, M.L., Hans, V., Pachman, L.M., and Fuleihan, R.L. (2019). Rituximab-associated Hypogammaglobulinemia in pediatric patients with autoimmune diseases. Pediatr Rheumatol Online J 17, 61. 10.1186/s12969-019-0365-y.
2. Liu, Y., Liu, Z., Yang, Y., Cui, J., Sun, J., and Liu, Y. (2023). The prognostic and biology of tumour-infiltrating lymphocytes in the immunotherapy of cancer. Br J Cancer 129, 1041-1049. 10.1038/s41416-023-02321-y.
3. Nahmad, A.D., Lazzarotto, C.R., Zelikson, N., Kustin, T., Tenuta, M., Huang, D., Reuveni, I., Nataf, D., Raviv, Y., Horovitz-Fried, M., et al. (2022). In vivo engineered B cells secrete high titers of broadly neutralizing anti-HIV antibodies in mice. Nat Biotechnol 40, 1241-1249. 10.1038/s41587-022-01328-9.
4. Matsushita, T., Yanaba, K., Bouaziz, J.D., Fujimoto, M., and Tedder, T.F. (2008). Regulatory B cells inhibit EAE initiation in mice while other B cells promote disease progression. J Clin Invest 118, 3420-3430. 10.1172/JCI36030.
5. Guberman Bracha, M., Biber, G., Zelikson, N., Shavit, S., Avraham, R., Vagima, Y., Bublik, D.R., Katz, Y., Barzel, A., Klapper, L.N., et al. (2025). Mouse B cells engineered to express an anti-HPV antibody elicit anti-tumor T cell responses. Front Immunol 16, 1613879. 10.3389/fimmu.2025.1613879.
6. Page, A., Hubert, J., Fusil, F., and Cosset, F.L. (2021). Exploiting B Cell Transfer for Cancer Therapy: Engineered B Cells to Eradicate Tumors. Int J Mol Sci 22. 10.3390/ijms22189991.
7. Nahmad, A.D., Lazzarotto, C.R., Zelikson, N., Kustin, T., Tenuta, M., Huang, D., Reuveni, I., Nataf, D., Raviv, Y., Horovitz-Fried, M., et al. (2022). In vivo engineered B cells secrete high titers of broadly neutralizing anti-HIV antibodies in mice. Nat Biotechnol 40, 1241-1249. 10.1038/s41587-022-01328-9.
8. Moffett, H.F., Harms, C.K., Fitzpatrick, K.S., Tooley, M.R., Boonyaratanakornkit, J., and Taylor, J.J. (2019). B cells engineered to express pathogen-specific antibodies protect against infection. Sci Immunol 4. 10.1126/sciimmunol.aax0644.
9. Muller, F., Taubmann, J., Bucci, L., Wilhelm, A., Bergmann, C., Volkl, S., Aigner, M., Rothe, T., Minopoulou, I., Tur, C., et al. (2024). CD19 CAR T-Cell Therapy in Autoimmune Disease - A Case Series with Follow-up. N Engl J Med 390, 687-700. 10.1056/NEJMoa2308917.
10. Chen, D., Kakabadse, D., Fishman, S., Weinstein-Marom, H., Davies, J., Boldison, J., Thayer, T.C., Wen, L., Gross, G., and Wong, F.S. (2023). Novel engineered B lymphocytes targeting islet-specific T cells inhibit the development of type 1 diabetes in non-obese diabetic Scid mice. Front Immunol 14, 1227133. 10.3389/fimmu.2023.1227133.
11. Brase, J.C., Walter, R.F.H., Savchenko, A., Gusenleitner, D., Garrett, J., Schimming, T., Varaljai, R., Castelletti, D., Kim, J., Dakappagari, N., et al. (2021). Role of Tumor-Infiltrating B Cells in Clinical Outcome of Patients with Melanoma Treated With Dabrafenib Plus Trametinib. Clin Cancer Res 27, 4500-4510. 10.1158/1078-0432.CCR-20-3586.








